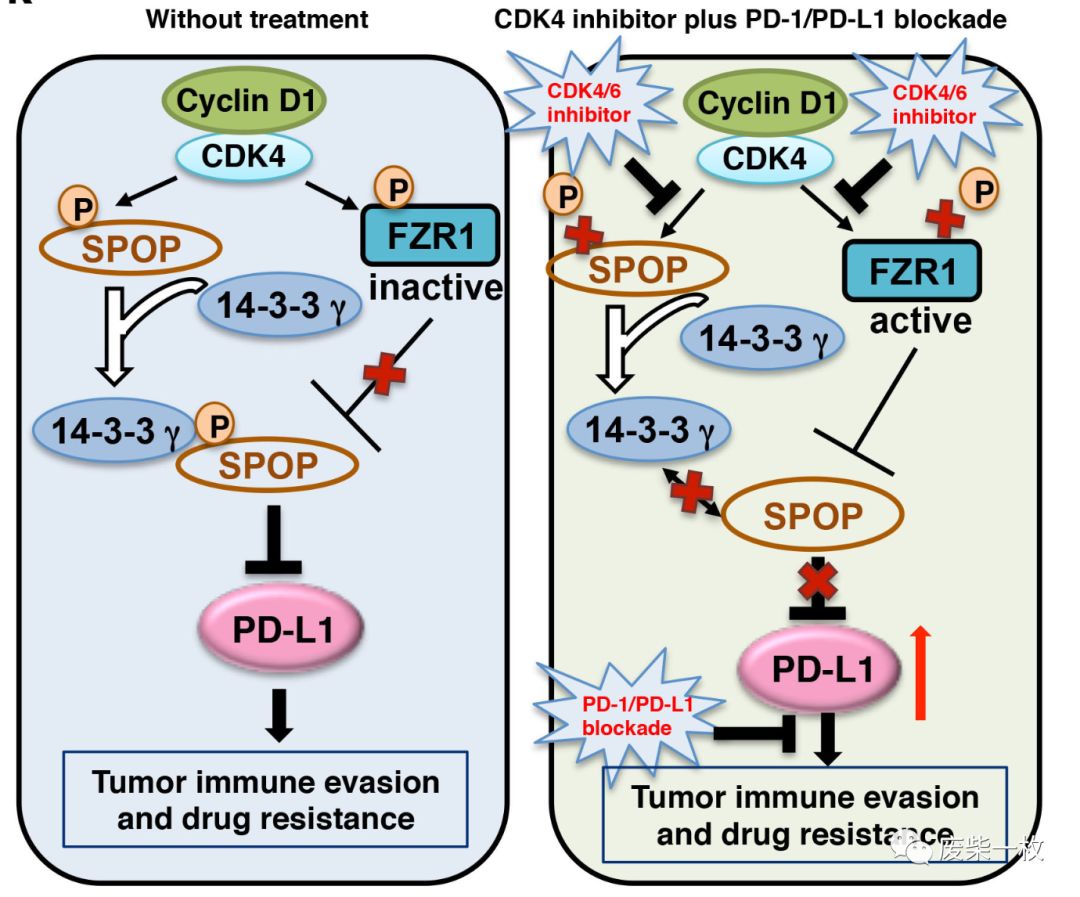
Cyclin D CDK4 kinase destabilizesPD-L1 via Cul3SPOP to control cancer immune surveillance
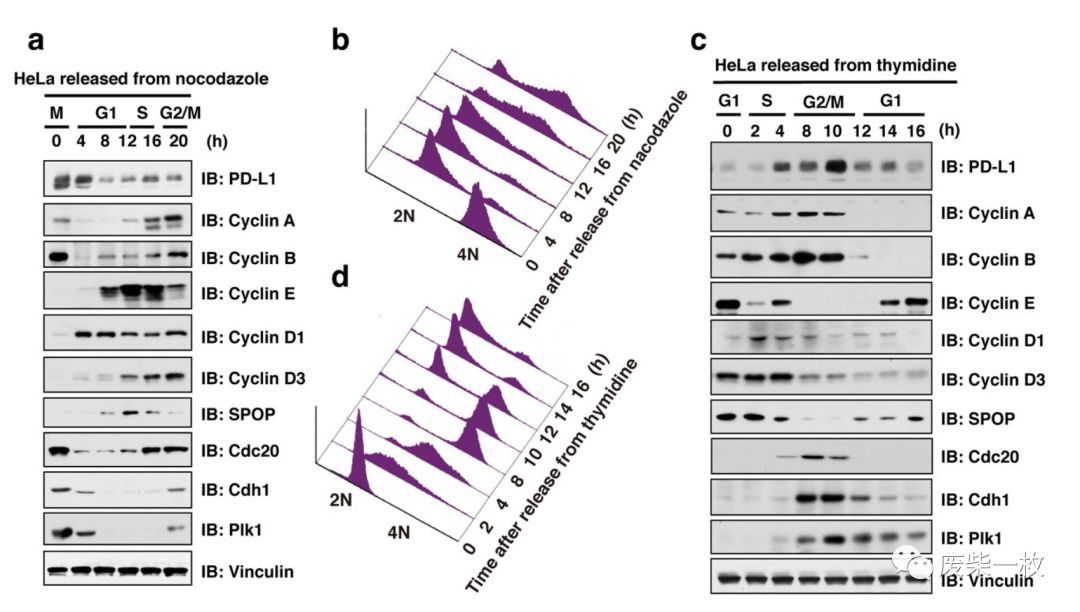
Figure 1: 用nocodazole将Hela细胞同步到M期,用thymidine将Hela细胞同步到G1/S期,使两者细胞同时从M期或G1/S期开始分裂增殖。取其同步之后不同时间点的细胞,Immunoblot分析发现细胞不同周期PD-L1的表达是处于波动状态。作者进而认为细胞周期与PD-L1的表达是有关系的。
细胞周期受细胞周期蛋白(cyclin)和细胞周期蛋白依赖性蛋白激酶(CDK)调控。目前发现有11种不同的cyclin,分别为A、B1、B2、C、D1、D2、D3、E、F、G和H。其中8种主要的cyclin己被分离。细胞周期蛋白可分为3类:S期周期蛋白,M期周期蛋白,G1期周期蛋白。S期周期蛋白为cyclin A,在S期开始表达,到中期时开始消失;M期周期蛋白为cyclin B,在S期开始表达,在G2/M期到达峰值,中期到后期转换时消失。G1期周期蛋白在脊椎动物中为cyclin C、D、E。
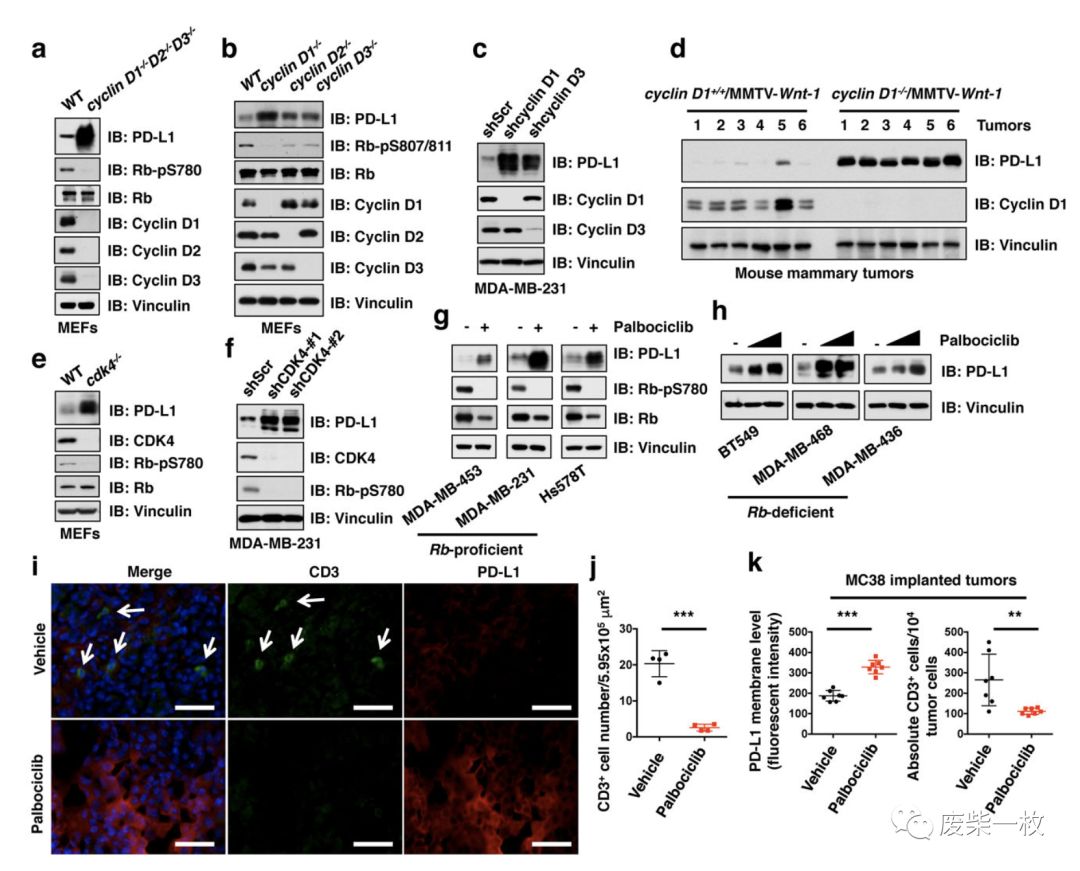
Figure2:图a-d说明,在MEF细胞中把CyclinD1/D2/D3敲掉,PD-L1的表达升高,出现了积累(而Figure S2a,b证明MEF细胞中敲掉CyclinA1/A2,
CyclinE1/E2,PD-L1的表达没有变化)。其中,a-c是在细胞中证明,d是在动物中证明。
哺乳动物细胞中,与G1期细胞周期蛋白结合的CDK激酶主要包括CDK2、CDK4和CDK6。cyclin D主要与CDK4和CDK6结合并调节后者活性;cyclin A常被划分为M期周期蛋白,但它也可与CDK2结合使后者表现激酶活性,说明cyclin A可能参与调控G1/S期转化过程。cyclin E在G1/S期与CDK2结合,呈现CDK2的激酶活性促进细胞进入S期。
图e-h说明在细胞中将CDK4敲除,PD-L1积累(Palbociclib是CDK4/6的抑制剂)
图i,j说明用MMTV-Erbb2建立肿瘤模型后,用Palbociclib抑制CDK4/6的作用,肿瘤组织中的PD-L1的表达增多,CD3阳性的T细胞数量减少。图K是用MC38建立模型后,得出相同的结论。
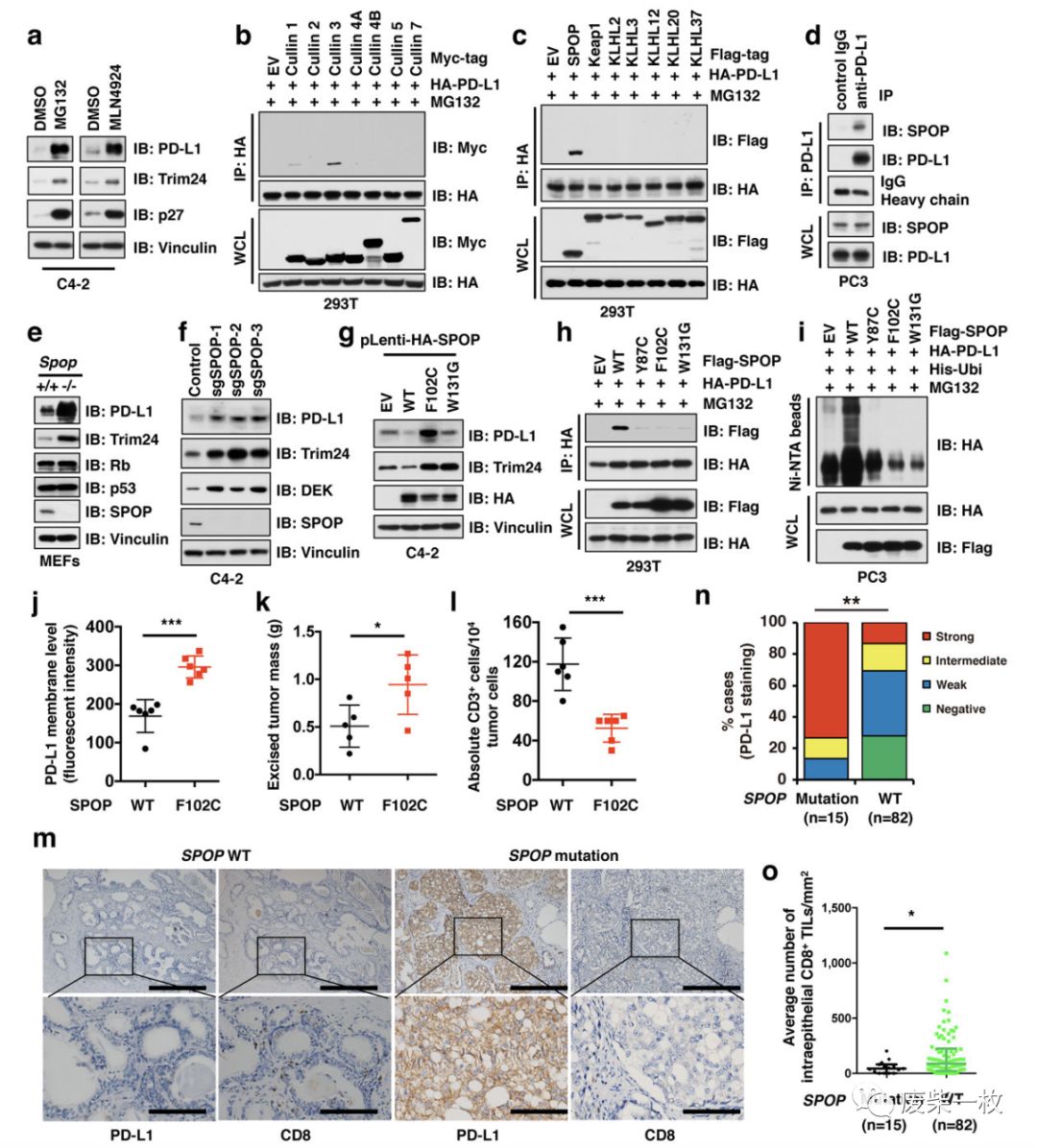
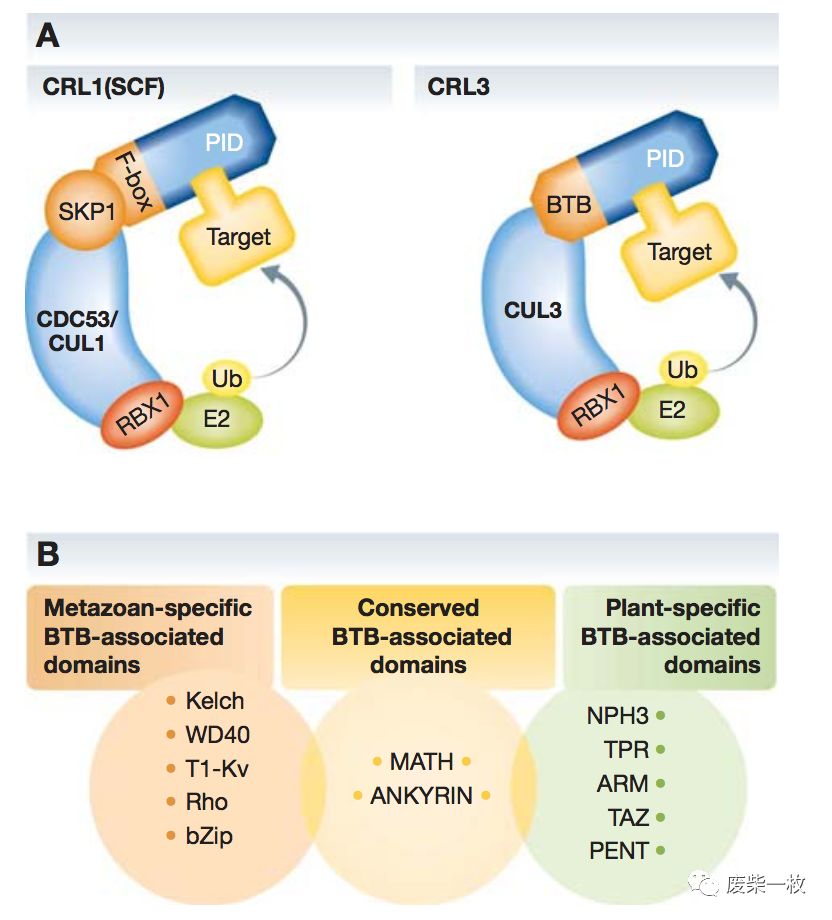
Figure3:作者认为PD-L1的积累可能与PD-L1的降解减少有关。蛋白的降解分为两类,一类是蛋白酶体的降解,一类是溶酶体的降解。MG132是蛋白酶体的抑制剂,MLN4924是基于Cullin的E3泛素连接酶的抑制剂。泛素化是一种重要的蛋白酶体降解途径。E3 泛素连接酶( E3 uibiquitin ligases) 是泛素 - 蛋白酶体系统中最大的一个 家族体系。到目前为止被发现的泛素连接酶已有 500 多种,它主要划分为三大亚家族: 第一类是拥有 HECT 结构域的 E3泛素连接酶; 第二类是拥有U.box结构域的泛素连接酶E3,另一类则是含有 RING 结构域的 E3 泛素连接 酶,由于它们都具有保守的 CUL 同源结构域,因此又被称 为 Cullin - Ring E3 泛素连接酶。Cullin蛋白主要划分为 Cullin1、2、3、4A、4B、5 和 7 7 个亚型。
Cullin - Ring E3 泛素连接酶家族成员以蛋白复合体的形式存在,包括小 RING蛋白( ROC1、ROC2) 、Cullin 蛋白、linker( 如 Skp1、Elongin B/ C、DDB1 等) 和底物识别亚基( 如 F - box、DCAF) 。Cullin 蛋白发挥支架的作用,其 N 端结构域结合接头蛋白和底物蛋白识别区域结合募集底物分子,C 端结构域则结合 RING 蛋 白( Rbx1 或 Rbx2) ,招募 E2 泛素结合酶。
a图说明PD-L1的积累与蛋白酶体的降解受到抑制有关,进一步说明与Cullin-E3泛素连接酶所介导的蛋白酶体降解受到抑制有关。
b图用cullin蛋白的7中亚型与PD-L1作IP分析,发现Cullin3与PD-L1结合(即向293T细胞中共转带HA标签的PD-L1质粒和带Myc标签的Cullin蛋白,收集细胞做IP,左侧IP:HA,是指用带HA的标签的磁珠去拉,右侧的IB:Myc是指用Myc的抗体去曝。)
c图用Cullin3的7种底物识别亚基与PD-L1作IP分析,发现SPOP与PD-L1蛋白结合(基于Cullin 3的E3泛素连接酶通过募集底物的adaptor蛋白识别其下游底物。如下图)。
a-c为外源性的IP,d图为内源性的IP。在PC3细胞中,用PD-L1的抗体去拉,lgG作阴性对照,发现在PC3细胞中,SPOP与PD-L1相互作用。
e-f说明将SPOP敲除,PD-L1蛋白积累。
g说明由于在某些细胞中,SPOP发生了突变,不能结合PD-L1,则PD-L1不能发生降解。
h是向293T细胞中共转入SPOP发生突变的质粒和PD-L1的质粒,IP分析显示,突变后的SPOP不能与PD-L1发生相互作用了。i图则为泛素化收到抑制。
a-i为细胞实验。在j-l图为动物模型。说明SPOP基因发生突变后,PD-L1积累增多,CD3阳性的T细胞降低,肿瘤变大。m-o为免疫组化证明SPOP发生突变的老鼠PD-L1和CD8T细胞表达增多。
The emerging family of CULLIN3-RING ubiquitin ligases (CRL3s): cellular functions and disease implications
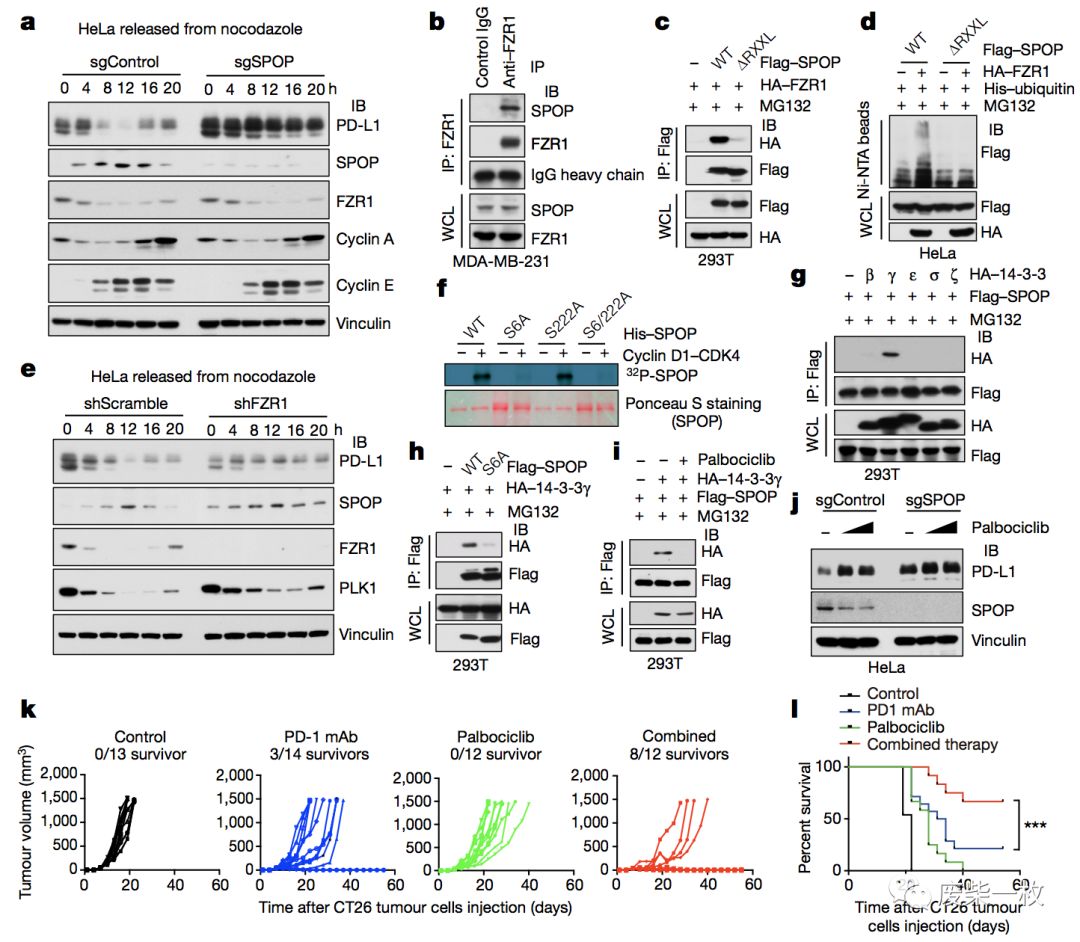
Figure 4 SPOP两个高度保守的丝氨酸-脯氨酸基序是Ser6和Ser222,图f说明cyclin D–CDK4磷酸化SPOP的位点是Ser6。
g,h图中作者发现14-3-3γ蛋白能与SPOP相互作用。
i,j是指将CDK 4/6抑制后(Palbociclib为CDK4/6抑制剂),spop与14-3-3γ不能发生相互作用。
a-j为细胞实验,k,i为建立动物模型后,抑制CDK4/6后,PD-L1积累,此时与抗PD-L1抗体联合使用后,荷瘤小鼠的生存率升高。
a为将SPOP基因敲除后,PD-L1积累。在control组,SPOP与FZR1的表达呈反相关。
b图为在MDA-MB-231细胞中,内源性的IP证明,SPOP与FZR1相互作用。
c图说明向293T细胞中共转入两种质粒后,FZR1与SPOP中的RXXL基序相互作用,将RXXL基序敲除后,FZR1与SPOP则不能发生相互作用。
e图为敲掉FZR1后,SPOP表达稳定,随后导致细胞周期进程中PD-L1蛋白水平降低。